| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
 妈妈化疗的艰难时刻,倍瑞博营养液带
当母亲被确诊为肺腺癌四期的那一刻,我们全家如坠冰窟。随之而来的化疗更是煎熬——
妈妈化疗的艰难时刻,倍瑞博营养液带
当母亲被确诊为肺腺癌四期的那一刻,我们全家如坠冰窟。随之而来的化疗更是煎熬——
 希望给没有阅读过的人一些体会
本文来自于此书,只是做一些摘抄,分享给深陷迷茫的人。文中观点真实与否,效果
希望给没有阅读过的人一些体会
本文来自于此书,只是做一些摘抄,分享给深陷迷茫的人。文中观点真实与否,效果
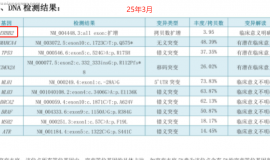
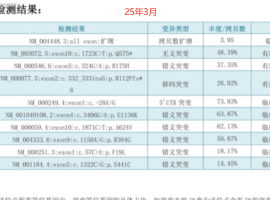 无靶点 her2扩增 二线白紫耐药后怎么
家父今年62岁,22年11月份颈部淋巴结穿刺,确诊肺低分化腺癌,分期是TxN3M1,基因检测
无靶点 her2扩增 二线白紫耐药后怎么
家父今年62岁,22年11月份颈部淋巴结穿刺,确诊肺低分化腺癌,分期是TxN3M1,基因检测
 父亲肺鳞癌,cT3N0M1a,ⅣA,脑转移
父亲2025年6月中旬因为头痛,一侧肢体不灵活入院,做了增强核磁,发现右侧额叶占位(3
父亲肺鳞癌,cT3N0M1a,ⅣA,脑转移
父亲2025年6月中旬因为头痛,一侧肢体不灵活入院,做了增强核磁,发现右侧额叶占位(3
 120万一针的抗癌药在肿瘤患者体内就
作者:Tony
一提起CAR-T,很多人都会想到的是其120万一针的恐怖价格,它是治愈癌症的
120万一针的抗癌药在肿瘤患者体内就
作者:Tony
一提起CAR-T,很多人都会想到的是其120万一针的恐怖价格,它是治愈癌症的





 寄予极大期望的易要不要再坚持吃段时间?才7天,可是感觉每天都在加重病情。明明,免疫组化是中阳啊!为什么要去测该死的突变,导致犹豫着始终不敢用。。是扩散太多了吗?失控了吗?感觉左眼周似乎有小的丘疹出现?加量吃么?还是换299804+184?
寄予极大期望的易要不要再坚持吃段时间?才7天,可是感觉每天都在加重病情。明明,免疫组化是中阳啊!为什么要去测该死的突变,导致犹豫着始终不敢用。。是扩散太多了吗?失控了吗?感觉左眼周似乎有小的丘疹出现?加量吃么?还是换299804+184? 显身卡
显身卡